ФАУНА МЛЕКОПИТАЮЩИХ ТЮМЕНСКОЙ ОБЛАСТИ
Современный видовой состав фауны млекопитающих
Периодизация голоцена, принятая нами,
включает два периода: доисторический (до XIV века н.э.) и исторический (после XIV века), включающий и современный период (с начала XX столетия). Видовой состав
фауны млекопитающих Тюменской области в исторический
и доисторический период, а также
характер распределения видов по зоогеографическим зонам и подзонам
представлены в базе данных. Всего в фаунистическом списке для
этого исторического периода представлено 98 видов (Гашев, 2001 б), но поскольку
на исследуемой территории некоторые виды животных из этого списка за последнее
столетие не встречались, а численность их в сопредельных районах существенно
сократилась, не оставляя надежд на существенное расширение ареала и появление в
пределах области, мы считаем их «вероятно исчезнувшими». К числу таких
видов относятся степная пищуха, степной сурок (Бибиков,
1989), малый суслик (Рузский, 1897), несмотря на то, что последний расширяет
свой ареал, в том числе и в северном направлении (Исмагилов, 1974), тарбаганчик (Рузский, 1897; Каталог млекопитающих СССР,
1981). И все же мы считаем нужным включить их в
фаунистический список именно как пример видов, исчезнувших в историческое время.
В.Л.Динец и
Е.В.Ротшильд (1996) без ссылок на конкретные места встреч распространяют на
часть территории Тюменской области ареалы таких видов, как рыжая вечерница (Nyctalus
noctula) и малая белозубка (Crocidura
suaveolens). Учитывая, что эти виды в
исторический период на территории области никем не отмечались, а карты в работе
названных выше авторов слишком мелки, что привело к искажению ареалов и
некоторых других видов, мы считаем невозможным включение этих трех видов в
фаунистический список Тюменской области. Не включаем мы в
современный фаунистический список пятнистого оленя (Cervus nippon), яка (Bos mutus) и
первобытного бизона (Bison priscus), разводимых в
вольерах на территории Тюменской области. Последнего в 1998 году завезли
из Канады на Полярный Урал для реакклиматизации,
однако, этих бизонов ряд систематиков рассматривает как подвид современного
американского Bison bison
L., 1758.
Таким образом, в настоящее время на
территории области могут быть встречены 94 вида млекопитающих из 8 отрядов,
которые и составляют современный фаунистический список млекопитающих Тюменской
области. Более трети всех видов составляют грызуны, второе и третье места
принадлежат соответственно хищным и насекомоядным
(Рис. 1).

Рис. 1.
Доля представителей отдельных отрядов млекопитающих
в териофауне Тюменской области.
Аннотированные списки видов всех
млекопитающих Тюменской области приведены в Приложении 1.
Четвертичная фауна млекопитающих
В силу ряда причин:
некоторая диспропорция палеонтологической изученности территории Тюменской
области - ее южные районы охарактеризованы несравнимо более подробно, чем
северные, в том числе и из-за ямальской трансгрессии
и ряда последовательных наступлений ледников в северной части территории, а
также существенная неравномерность в изучении фаун отдельных эпох плейстоцена,
- последовательная реконструкция
пространственно - временной динамики ареалов млекопитающих Западной Сибири
представляется, по мнению Н.Г.Смирнова (1988), пока невозможной.
Поэтому мы не останавливаемся подробно на доисторическом
этапе формирования териофауны области, а сошлемся на
ряд специальных работ, посвященных этой
проблеме (Малеева, 1970; Смирнов, 1976; Зажигин,
1980; Смирнов, Большаков, Бородин, 1986; Бородин, 1988; Малеева, Стефановский, 1988 и др.), обозначив лишь основные этапы
формирования териофауны области и приведя перечень
основных видов, обитавших на территории Тюменской области на рубеже
плейстоцена-голоцена.
В формировании териофауны
Западной Сибири Н.Г.Смирнов с соавторами (1986) выделяют два основных этапа:
1) плейстоценовый - в это время южные границы тундрового
комплекса млекопитающих доходили до 640 с.ш.,
а южнее располагались тундро-степи и степи;
2) послеплейстоценовый (голоценовый)
- период образования таежно-лесного комплекса, а П.В.Косинцев
(1999), кроме того, указывает на автохтонную и миграционную составляющие
процесса формирования териофауны севера Западной
Сибири. При этом, применительно к голоценовой истории
фаунистических комплексов остро стоит вопрос: на каком из этапов развития голоценовые сообщества утратили те или иные черты,
унаследованные от плейстоценовых экосистем (Смирнов и др., 1999).
Обобщая различные литературные данные (Пидопличко, 1951; Каталог млекопитающих..., 1981; Малеева,
1983; Смирнов, Большаков, Бородин, 1986; Малеева, Стефановский,
1988; Природа Тюменского..., 1991; Косинцев,
Ситников, 1998, Косинцев, 2001, Воробьев, 2001 и
др.), можно представить примерный список плейстоценовых млекопитающих Тюменской
области (Приложение 1).
Фактические подтверждения обитания ряда
видов из такого списка в настоящее время отсутствуют, но находки их в
сопредельных регионах позволяют считать вероятным их встречи в исследуемом
регионе.
Формирование современной фауны млекопитающих
По представленности
видов млекопитающих, обитающих на Западно-Сибирской равнине и примыкающей к ней
арктической акватории в современности, Тюменская область занимает первое место
в регионе - 89 % всех млекопитающих. Но в целом фауну Западной Сибири можно
считать достаточно бедной, особенно по сравнению с фауной Восточной Сибири.
Основной причиной бедности териофауны Западной Сибири
являются последствия ямальской трансгрессии,
покрывшей весь север водами Бореального бассейна, и последующих плейстоценовых
оледенений, самое мощное из которых - самаровское,
охватило более половины суши исследуемой территории, создав дополнительно
значительный подпрудный бассейн на р.Оби (Старков, Тюлькова, 1996), а также относительная отдаленность горных рефугиумов, питающих миграционный поток в голоцене. Теплолюбивая фауна была существенно обеднена во время четвертичных
оледенений, когда на территории области распространились тундровые,
лесотундровые и перигляциальные ассоциации, а
современное животное население стало формироваться после исчезновения ледников
и с формированием таежно-лесного комплекса в позднеплейстоценовое время и в
голоцене, однако, нет оснований говорить о чередовании фаунистических
комплексов ледниковых и межледниковых эпох (Смирнов, Большаков, Бородин, 1986),
шла лишь их трансформация на конкретных территориях
за счет включения или выпадения некоторых видов в результате сдвига ареалов в
северном или южном направлениях вслед за смещением границ
природно-климатических зон. Причем, наряду с автохтонами шло заселение Западной
Сибири и с запада, и с востока (этот процесс продолжается и в настоящее время).
Именно поэтому фауна Западной Сибири вообще (Шварц, 1989) и фауна млекопитающих
Тюменской области, в частности, имеют смешанный европейско-восточносибирский
характер, представляя комплекс видов европейских бореальных лесов, ряда видов
восточносибирского происхождения и североатлантических видов. Процесс
проникновения видов на территорию Тюменской области с востока и с запада
продолжается и в настоящее время, а благодаря все большему освоению этой территории человеком, наметился
вектор заселения европейскими видами и более северных территорий (обыкновенная
полевка, рыжая полевка и др.). По числу видов в Тюменской области доминируют
представители европейского типа фауны, кроме северной тайги, где их сменяют
сибирские виды на общем фоне транспалеарктического
ранга, а в Субарктике эти особенности фауны ослабевают, уступая место
циркумполярным закономерностям. Видов-эндемиков в териофауне
Тюменской области нет. Преобладание европейского компонента в целом по области
мы объясняем наличием опережения по времени наступления и отступления ледников
в Евразии в западных районах по сравнению с восточными
(Пузанов, 1938).
В целом закономерности распределения
видов териофауны Тюменской области в исторический и
современный периоды по эколого-географическим признакам видны из таблицы 1.
Таблица 1.
Распределение видов териофауны Тюменской области по эколого-
географическим признакам
|
Космополиты 3 |
Циркум- полярные и бореальные |
Евро- Азиатские |
Европейские |
Азиатские |
Азиатско- Американ-ские * |
|
Полярные |
11 |
- |
1 |
1 |
1 |
|
Бореальные |
10 |
11 |
23 |
13 |
4 |
|
Температно- аридные |
- |
12 |
5 |
2 |
- |
Примечание: * -
сюда включены и чисто американские виды.
Необходимо отметить и такой компонент
фаунистического комплекса, как виды, целенаправленно акклиматизированные
человеком и расселившиеся на исследуемой территории (выхухоль, ондатра,
европейский подвид речного бобра, енотовидная собака и американская норка), а
также виды, восстановленные человеком после их исчезновения в доисторическое
(овцебык, благородный олень) и историческое время (кабан). Виды-эвсинантропы (домовая мышь и
серая крыса) в своем распространении в Тюменской области также связаны с
деятельностью человека, его жильем, хотя в летнее время и выселяются в природу.
Особняком стоят домашние животные,
существование которых целиком зависит от человека. Так еще в среднем голоцене на территории
Западной Сибири (в основном в ее южной части) были интродуцированы
четыре новых вида - крупный рогатый скот, домашняя лошадь, овца и
коза (Косинцев,
1988). Два первых
вида из них имели природные
аналоги в териофауне
лесостепной и степной зон - тура и дикую лошадь, два же последних вида
не имели аналогов в аборигенной фауне. Повлияла ли эта интродукция на териофауну и если повлияла, то как, - пока не ясно (Косинцев, 1988). Ископаемые останки собаки тоже встречается
в тех же месторождениях голоценовой териофауны. Практика показывает, что этот вид в условиях
юга области и даже таежной зоны может существовать относительно самостоятельно.
Так, например, имеются сообщения об образовании устойчивых стай одичавших собак
не только в южной Европе (Бараташвили, 1990), но и в Западной Сибири (Телегин,
Ивлева, 1990). В последнее время поток интродуцентов
на территорию области сильно возрос - это свиньи, кошки, кролики, нутрии из
млекопитающих и целый ряд экзотических видов из разных систематических групп
позвоночных. Так, известный тюменский таксидермист В.Е.Кюкен
рассказывал об обитании в один из 60-х годов в низовьях р.Вагай сбежавшего из плавучего зоопарка крокодила,
который дожил до осени; из птиц известны случаи обитания летом в г.Тюмени
канареек; из млекопитающих - макаки резус, сбежавшей со станции юных
натуралистов и т.д. Безусловно, большинство из них не могут существовать в
естественных условиях без участия человека и по этой причине не включаются нами
в фаунистический список Тюменской области.
Все сказанное выше можно проиллюстрировать на
динамике видового состава млекопитающих за последний исторический период, в
качестве временных вех выбрав рубеж 18 века и 21 столетия (Трансформация фауны позвоночных в
исторический период). Первая связана с периодом массового проникновения
русских служилых людей на территорию нынешней Тюменской области, а вторая - с
последним, наиболее интенсивным этапом ее промышленного освоения. Конечно, конкретные данные о видовом составе животных на рубеже
16-17 веков скудны, но имеющиеся, а также ретроспективные данные, учитывающие
не столько данные о наличие отдельных видов (основной облик териофауны
региона сложился уже к середине голоцена), сколько известные факты появления
или исчезновения тех или иных видов, позволяют воссоздать вполне, на наш
взгляд, репрезентативную картину. Общий фаунистический список
позвоночных животных Тюменской области (без учета человека и домашних
животных), составленный нами для указанного исторического периода, включает в
себя 557 видов позвоночных животных и в т.ч. - 98 видов млекопитающих. Распределение млекопитающих на рубеже
16-17 веках и в начале 21 века приводится в таблице 2.
Конечно же, нужно иметь в виду, что
изменения в фауне позвоночных происходили не только в силу действия
антропогенных факторов: сказалось и изменение глобального климата, и
естественные процессы расселения видов (Западная Сибирь с послеледниковых
времен постепенно заселяется европейскими, центрально-азиатскими и
восточносибирскими видами). Однако, не вызывает сомнения, что антропогенная
трансформация ландшафта способствовала расселению одних видов и сокращала
ареалы других.
Таблица 2.
Фауна позвоночных
животных территории Тюменской области
|
Классы позвоночных |
Начало
17 века |
Начало
21 века |
Исчезло видов |
Появилось видов |
Разница |
|
Млекопитающие |
86 |
89 |
8 |
11 |
+3 |
|
Всего позвоночных: |
518 |
552 |
12 |
46 |
+34 |
В отношении таких видов позвоночных, как
большой сурок, дрофа, стрепет, авдотка, исчезнувших с описываемой территории,
можно смело констатировать прямое влияние хозяйственной деятельности человека.
Оно же способствовало проникновению на эту территорию краснощекого суслика,
зайца-русака, сизого голубя и многих других. Целый ряд видов
был завезен самим человеком целенаправленно (ондатра, енотовидная собака,
американская норка, овцебык, благородный олень, карп, судак, толстолобик, белый
амур и др.) или случайно (мышь домовая, крыса серая, озерная лягушка, ротан, верховка, уклейка, и
т.д.).
Из таблицы 2 видно, что за четыреста лет в целом с территории нынешней Тюменской области исчезло лишь 7 видов позвоночных и из них 4 вида млекопитающих (хотя численность многих сохранившихся резко сократилась и судьба этих видов вызывает тревогу !), в то же время появилось 40 новых видов, 11 из которых - млекопитающие. Кстати, подобные закономерности отмечает и К.В.Граждан (2002) в своей работе по населению птиц Северо-Восточного Алтая за последние 40 лет (из 213 видов птиц 14 видов встречены только в 60-е годы, а 32 вида – только в конце 90-х). Таким образом, широко муссируемый тезис об обеднении видового состава животного мира под воздействием антропогенных факторов не выдерживает критики. Однако, сам факт существенной трансформации видового состава является показателем наличия антропогенного воздействия (прямого или опосредованного) на фауну, а степень этой трансформации - показателем силы этого воздействия, и в этом смысле данные показатели обязательно должны использоваться при оценке экологического состояния тех или иных территорий в системе экологического мониторинга.
Анализ же качественных изменений в
видовом составе (изменение доли синантропов, антропофилов
и антропофобов) помогает вынести суждение о
направленности изменений среды обитания, составить прогноз возможных дальнейших
изменений в фауне. В этом плане современная фауна позвоночных животных
Тюменской области, в целом, и млекопитающих, в частности, значительно
обогатилась синантропами и антропофилами, что,
кстати, повышает общий уровень ее устойчивости к антропогенным факторам.
Однако, при всей важности исторического
аспекта биоразнообразие фауны материковых регионов
определяется и современным ей состоянием экосистем и климата (Гынгазов, Миловидов, 1977), испытывая существенные
колебания в ходе различных климатических эпох голоцена, когда относительно
прохладные и влажные периоды сменяются многовековыми
засушливыми и теплыми эпохами. Всего в течение голоцена отмечено шесть
таких крупных изменений. Смены этих эпох влекли и влекут за собой циклические
колебания ареалов многих западносибирских видов, в том числе и млекопитающих, в
северном или южном направлении с последующим возвращением в исходное положение.
Вообще же в силу своей географической открытости с севера и юга в сочетании с равнинностью Западная Сибирь имеет четкую арктико-континентальную детерминацию, что и обуславливает
видовое разнообразие различных природных и зоогеографических зон и подзон Тюменской области.
Географические закономерности в фауне млекопитающих области
Териофауна области крайне бедна на севере и значительно
обогащается к югу (Рис.2), при этом проявляется дифференциация по широте и
более слабая (связанная с продвижением европейских и восточносибирских видов) -
по долготе. В Субарктике ареалы животных вообще носят циркумполярный характер,
в зоне тайги это проявляется реже, а еще южнее отсутствует вовсе. Нарастание
видового богатства млекопитающих с продвижением к югу наблюдается вплоть до
южных границ области (подзона средней лесостепи).
Анализ материала показал, что описанная картина не меняется и с учетом видов,
возможно исчезнувших в течение последнего столетия, что позволяет предположить,
что причиной такого распределения видов являются какие-то естественные, а не
антропогенные факторы. Увеличение видового богатства млекопитающих при
продвижении с севера на юг в пределах Тюменской области отмечается и в горных
районах Урала. Отметим, что нахождение максимума видового богатства
млекопитающих в целом (Лаптев, 1958) и мелких млекопитающих (Равкин и др., 1996) в зонах подтайги
и лесостепи отмечено для всей Западной Сибири.
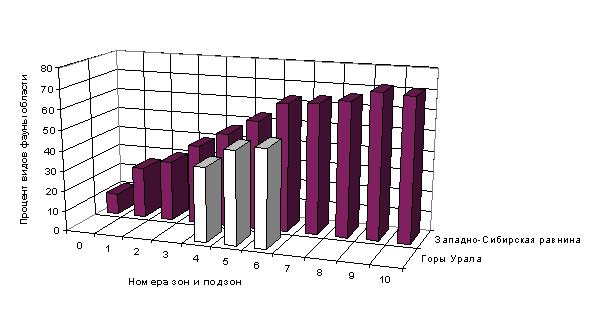
Рис. 2. Распределение видов
млекопитающих по зоогеографическим зонам
и подзонам
(номера зон и подзон см. в соответствующих разделах сайта).
Очевидно, что значение отдельных
видов млекопитающих, включенных в формальный список териофауны
Тюменской области, неодинаково. В первую очередь, это связано с количественной представленностью
тех или иных видов. Для териофауны описываемого
региона характерна высокая степень доминирования на всех трофических уровнях, а
абсолютными доминантами по численности являются грызуны и насекомоядные. В зоне тундр такими доминантами являются тундряная
бурозубка из насекомоядных и сибирский лемминг из грызунов (субдоминанты -
копытный лемминг и полевка Миддендорфа); в лесотундре
- возможно, полевка-экономка; в таежной зоне доминирующая роль среди
насекомоядных (и всех млекопитающих) с продвижением на юг переходит к
обыкновенной бурозубке, а среди грызунов - к красной полевке (полевка-экономка
и средняя бурозубка занимают субдоминирующее
положение); в подтайге
возрастает роль рыжей полевки, которая наряду с полевкой-экономкой прочно занимает субдоминирующее
положение (а в западной части зоны даже выходит в доминанты среди грызунов),
обыкновенная бурозубка сохраняет роль абсолютного
доминанта; в лесостепной зоне на первое место среди грызунов выходят
узкочерепная полевка, полевка-экономка и полевая мышь (в разные годы - один из
видов), субдоминирующее положение занимают красная
полевка, обыкновенная полевка и лесная мышь, среди
насекомоядных лидирующее положение сохраняется за обыкновенной бурозубкой. В
горных районах Урала доминантами являются красная и
красно-серая полевки из грызунов (в разных высотных поясах) и тундряная и обыкновенная бурозубки из насекомоядных (в
северных и южных районах, а также в тундровой и горно-таежной подзонах соответственно).
Из мелких млекопитающих шире всего по числу заселяемых широтных зон и подзон (в лесной части) отмечены тундряная
и малая бурозубки, домовая мышь, полевки: экономка, красная и водяная - они
встречаются повсеместно (Равкин и др., 1996). Из
других групп млекопитающих сюда нужно отнести горностая, ласку, лисицу, волка и
лося. Больше всего видов мелких млекопитающих предпочитает, судя по
максимальному среднему обилию, подтаежные леса (Равкин
и др., 1996). Наряду с зональной проявляется и
провинциальная изменчивость распределения млекопитающих. Так, преимущественно в западной части области
обитают (особенно на севере) европейский крот, полевка Миддендорфа,
лесной лемминг, лесная куница, европейская норка и ряд других видов, а в
восточной части области - крот сибирский и бурозубка бурая. Хотя бы в одной из подзон
увеличивается обилие к западу у обыкновенной бурозубки, куторы,
лесной мыши, рыжей полевки, а противоположная тенденция отмечена для равнозубой, тундряной, бурой,
средней, малой и крошечной бурозубок, красной, водяной и темной полевок (Равкин и др., 1996). Вообще же Б.С.Юдин (1988) отмечал
снижение численности к востоку у западно-палеарктических видов и обратную
тенденцию у восточно-палеарктических.
В целом, численность мелких млекопитающих
в Западной Сибири на 31 % определяется зонально-подзональными
особенностями, на 22 % - облесенностью, на 19 % -
составом древостоя и лишь на 2 % антропогенным влиянием (Равкин
и др., 2002).
Динамика численности млекопитающих
Изучение динамики численности животных само по себе, уже по определению,
является мониторингом. Однако,
принимая как аксиому
тот факт, что
динамика численности наряду с популяционными механизмами саморегуляции зависит от условий окружающей среды, мы можем
рассматривать особенности ее в качестве критерия оценки этой среды,
подверженной трансформациям под действием естественных или антропогенных
факторов. В этом смысле динамика численности животных (годичные флуктуации (Одум, 1986)) является важным объектом экологического мониторинга не только в прикладном (для
охотоведения, сельского хозяйства и т.д.), но и в теоретическом плане. В настоящей работе мы на основе собственных многолетних данных и
анализе литературного (Балахонов и др., 1997 и др.) и ведомственного (Фонд...,
1992; Обзор..., 1995, 1996, 1997, 1998, 1999; Ресурсы..., 1996) материала по
динамике численности млекопитающих Тюменской области постараемся осветить
некоторые особенности использования этого показателя в системе экологического
мониторинга (Гашев, 2000 д). Прежде всего,
нужно иметь в виду, что годовые и
многолетние колебания численности млекопитающих во многом определяются
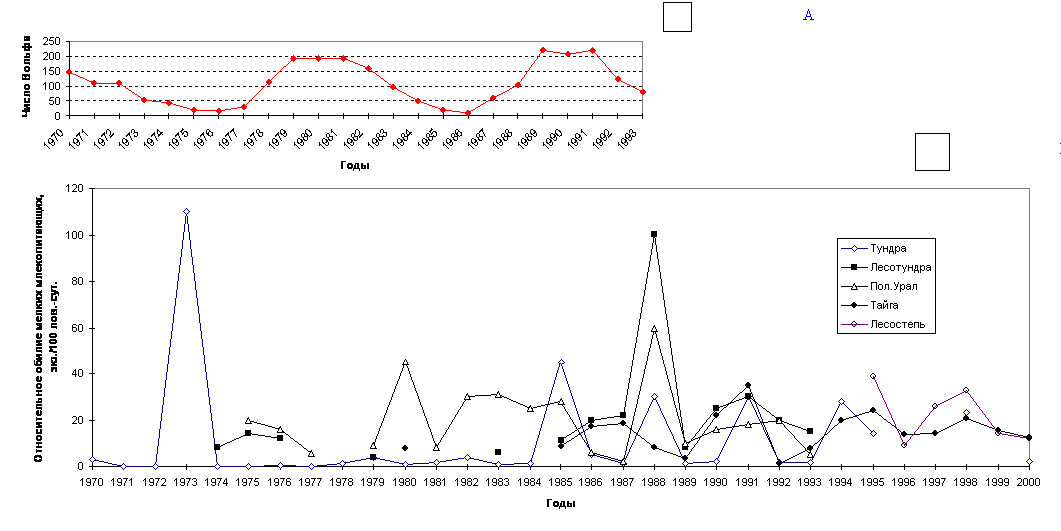
естественными причинами. Так, например, В.С.Дьячков (1989), С.А.Корытин
(1994) и др. опосредованно связывают их с 11-летними циклами солнечной
активности (Рис. 3 а). Поэтому далеко не всегда снижение или, наоборот, рост численности
того или иного вида может свидетельствовать, скажем, о каком-то действии
антропогенных факторов. В этом плане хорошо известен пример изменения
численности зайца-беляка и связанные с ними флуктуации рыси в Канаде во второй
половине 18 - первой половине 20 века (MacLulich, 1937). Колебания численности
зайца-беляка имели достаточно четкую 10-летнюю периодичность (в среднем - 9.6
года), и этим колебаниям соответствовали колебания численности рыси, питающейся
зайцем-беляком. При этом пики численности зайца на год или чуть более
предшествовали пикам численности рыси. Такие же регулярные естественные
колебания численности характерны и для других животных. При этом многолетние
флуктуации численности отдельных видов условно делят на цикличные и не цикличные,
используя индекс цикличности популяций - s (Уильямсон, 1975 цит. по Кривошеев, Кривошеева, 1998). Мы
предлагаем использовать интегральный материал по всей группе мелких
млекопитающих. Так, численность мелких млекопитающих значительно колеблется по
годам: в Тюменской области эти флуктуации прослежены более чем за 30 лет: с
1970 по 2002 гг. (Рис.14 б)(Балахонов и др., 1997,
наши данные и др.). Анализируя картину в целом, можно отметить, что за этот
период отмечалось 10 депрессий (1971-72; 1974; 1977-78; 1981; 1984; 1986-87;
1989; 1992-93; 1996; 1999-2000) и 10 пиков численности (1970; 1973; 1975-76;
1979-80; 1982-83; 1985; 1988; 1991; 1994-95; 1998) мелких млекопитающих.
Периодичность колебаний составила около 3 лет (в среднем 2.9 года). Хорошо видно, что, несмотря на некоторые
смещения сроков пиков и спадов в разных природных
зонах, общая картина сохраняется везде. Бросается в глаза, что в наиболее
экстремальных условиях среды (тундра и лесотундра) выраженность колебаний выше
по амплитуде. Причем, говоря о более экстремальных условиях среды в тундре и
лесотундре, мы имеем в виду объективные характеристики этих условий, соглашаясь
с тем, что виды, населяющие эти природные подзоны,
безусловно, имеют целый ряд адаптивных приспособлений для переживания этих экстремальных
условий. Сама же «экстремальность» отражена в значениях коэффициентов
«вязкости» (К) и «упругости» (G) среды в
нашей модели устойчивости. Образно говоря, в этих природных зонах под одним и
тем же «шаром» (сообществом) плоскость прогибается незначительно, а сама
поверхность «чаши» практически лишена шероховатости. В этих условиях маятникообразные колебания в системе, безусловно, больше по
амплитуде. Это и наблюдается в отношении колебания численности мелких
млекопитающих. При
анализе этого материала мы отмечаем, что еще более правильную периодичность
имеют не пики или депрессии численности мелких млекопитающих, а точки перехода
кривой динамики численности через среднюю многолетнюю - это происходит строго
через 2 года на третий. Выявленная закономерность, безусловно, может
быть использована при прогнозировании вспышек численности мелких млекопитающих
при проведении ежегодных контрольных учетов в рамках экологического
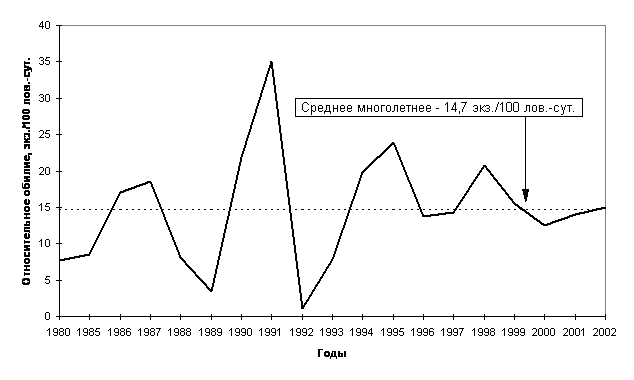
мониторинга. В таежной зоне Тюменской
области за период с 1985 по 2002 год отмечено 4 пика численности и 5 депрессий
(Рис. 4), то же самое отмечают Н.С.Москвитина с соавторами (2000) для лесных
полевок в Томском Приобье.
Причем, флуктуации численности насекомоядных и грызунов, по нашим данным, идут
синхронно, хотя, по мнению ряда авторов (Бачурин и др., 1975), эти колебания у
разных видов мелких млекопитающих могут не совпадать, и тогда состав доминантов по годам будет меняться. Так
летом 2002 года при невысокой численности грызунов отмечалось высокое обилие
насекомоядных, обеспечившее некоторый рост численности мелких млекопитающих в
целом. Динамика численности мелких млекопитающих в разных биогеоценозах
близка, хотя в ряде из них, играющих роль «стаций переживания» (в Среднем Приобье это - спелые зеленомошно-кустарничковые
кедровники), численность меняется не так значительно, оставаясь достаточно
высокой даже в годы депрессий.
При попытке установить
корреляционные зависимости численности красной полевки и обыкновенной бурозубки
с различными морфофизиологическими
характеристиками в предшествующий вегетационный сезон наиболее сильная
положительная связь отмечена с индексом печени
самок-сеголеток (r = 0.72 ± 0.40 и 0.91 ± 0.24 при P < 0.05
соответственно для каждого вида).
Корреляционной связи обилия названных выше доминирующих в таежной зоне
видов с успешностью их размножения или с солнечной активностью (в предыдущий
сезон) в 1987-97 гг. нами не установлено.
Зная естественные закономерности динамики численности тех или иных видов
млекопитающих в определенных регионах, по наблюдаемым отклонениям от них можно
выносить суждения о влиянии каких-то возмущающих факторов на популяцию вида,
что чрезвычайно важно в системе экологического мониторинга. Примером таких
наблюдений являются наши исследования влияния нефтедобычи (в первую очередь,
нефтяного загрязнения) на мелких млекопитающих в Среднем Приобье
Тюменской области в 1987-1991 гг.
Нужно отметить, что биогеоценотическая
роль ряда видов из формального списка минимальна в силу их естественной
невысокой плотности популяций (летяга, крошечная бурозубка, тушканчик большой и
др.), либо в силу «случайного» характера пребывания на территории Тюменской
области (к последним относятся представители категории
«возможно встречающихся» видов). Ценность таких видов для целей экологического
мониторинга ниже, однако, было бы методологически ошибочно исключать их из поля
зрения, так как они могут являться индикаторами изменения
например климатических условий существования на границе своего ареала. В фауне
млекопитающих Тюменской области установлены и сезонные изменения видового
состава, распространения и численности популяций видов. Так,
зимой отсутствуют все виды рукокрылых, широко к югу расселяется песец; на
севере таежной зоны сокращается численность лося и увеличивается численность северного
оленя, а на юге резко снижается количество косули; часть видов (бурый медведь,
бурундук, суслики и др.) впадает в спячку, и их роль в биогеоценозе также резко
снижается (Лаптев, 1958).